

Earstones of Mohawk Herring
Strontium geochemistry is a distinct tag for Mohawk fish
Alewife (Alosa pseudoharengus) and Blueback Herring (Alosa aestivalis) – collectively “River Herring” - like other North Atlantic anadromous fishes, currently occur at historically low levels. Thus an understanding of their reproduction, growth, and migration is important for conservation (Turner and Limburg, 2014). Recall that anadromous fish migrate from the ocean to fresh water to spawn, and the runs of these fish in the spring can be amazing. So the young are born in our rivers, then they leave, and with a little luck they return to spawn. Otoliths or earstones in these fish have a high fidelity to water chemistry, and the water in the Mohawk is distinctive enough - compared to the Hudson - to tag the homeland of these fish once they leave the Watershed.
A significant population of River Herring occurs in the Hudson watershed, but the Mohawk River population is interesting and curious because in order to thrive in this watershed they need to negotiate the complicated Waterford lock system (Locks E2 through E6 at Waterford with an elevation change of 169 ft). River Herring colonized the Mohawk after construction of the lock system nearly 200 years ago. Conventional migration up the natural river channel was not possible because the 90-foot Cohoes Falls near the confluence with the Hudson forms a natural ecological barrier. These fish are important forage for birds, mammals, and piscivorous fishes such as Striped Bass, among others.

The Blueback Herring (Alosa aestivalis) ranges from Nova Scotia to Florida, and is common in the Hudson and lower Mohawk. This drawing was prepared by Ellen Edmonson and Hugh Chrisp as part of the 1927-1940 New York Biological Survey project conducted by the Conservation Department (the predecessor to the NYS Department of Environmental Conservation). Permission for use of image is granted by NY State Department of Environmental Conservation. This Image (and topic) was featured in the 2016 Mohawk Watershed Symposium where Karin Limburg, highlighted below, was the keynote speaker..
An otolith or earstone is a calcium carbonate (aragonite) structure that is a critical part of the hearing and balance system in typical fish. An otolith is a rock (mineral actually), not bone, and continuous daily growth incorporates trace elements in water. So as the otolith grows it records the chemical chronology of the fish (Elsdon et al., 2008). Thus when a fish grows, seasonal changes in temperature and water chemistry are recorded in the otolith, and of course seasonal changes (winter and summer) are recorded in the annual growth rings, which are commonly used to estimate the age of a fish. Otoliths incorporate trace elements and isotopes from ambient water, and thus the compositional changes outward in the otolith record changes in water chemistry over time (see Campana 1999 for an overview and Turner and Limburg, 2015 and Limburg and Turner, 2016 for a Mohawk/Hudson specific overview).
Considerable effort has gone into using the geochemistry of otoliths to determine the provenance of migratory fish, and the primary approach has been to investigate those elements that are common in marine and fresh water, and those that can substitute into the aragonite structure of the otolith. Most studies focus on ratios of elements and isotopes: strontium/calcium (Sr/Ca), barium/calcium (Ba/Ca), isotopes of strontium (87Sr/86Sr), and stable isotopes of oxygen (Turner and Limburg, 2014). Provenance here is a term used to signify the source or origin – the same term is used for wine.
Because strontium can replace calcium, it can be found in bone, teeth, and otoliths. In terms of recording a clean strontium signal, bone is good, teeth are very good, and otoliths are excellent. There are several reasons for this, but otoliths have continuous outward growth and no elemental exchange with previously accreted layers, and this the not the case with bone.
Interesting and exciting research has emerged from Karin Limburg and Sara Turner, and most of this post is based on two papers they recently published (cited above, listed below). Early work in the Hudson and Mohawk showed that there is a slight difference in the Sr/Ca ratio between the Mohawk and the Hudson (Mohawk is higher), and that Sr/Ca is much higher than either river in the marine environment (Limburg et al., 2001). Some of that differences in the freshwater ratios may be attributed to watershed geology: the large fraction of limestones (strontium-rich) in the Mohawk and more abundant crystalline rocks in the upper Hudson.
Later work showed that the 87Sr/86Sr ratios are also significantly different between the two watersheds. Thus these isotopic ratios in the otoliths of migratory herring could be used as a sort of bar code that indicates where the fish lived, and for how long (see Turner and Limburg, 2014).
There are two aspects of strontium that are used in these analyses. Strontium (Sr2+) readily substitutes for calcium (Ca2+), so if Sr is available it can be taken up by organisms. The Sr/Ca ratio is very different in marine and freshwater otoliths because marine water has much higher concentrations of Sr, and therefore the substitution is greater when the fish lives in the ocean. This is slightly complicated when fish spend time in an estuary because the amount of available Sr varies depending on salinity: the marine Sr signal will be less distinct the farther up the estuary the fish lived (i.e. Albany v NYC). The other important aspect of strontium in the otolith is more involved and to explore this we need to get into isotopes.
Strontium isotopes
Strontium has four isotopes that have different natural abundances: 84Sr (0.56%), 86Sr (9.86%), 87Sr (7.0%) and 88Sr (82.58%). Of these isotopes only 87Sr can be radiogenic, which means that some of it is formed through the radioactive decay of a parent isotope, which in this case is 87Rb (acts like potassium). The other isotopes are stable and non-radiogenic (and thus do not change over time). The ratio of 87Sr/86Sr is commonly used in geology because there are a number of distinctive crustal processes that allow the ratio to change but the most important from the perspective of otolith growth are chemical composition of bedrock and age of bedrock.
Because isotopes of Sr don’t fractionate to any significant degree during biological uptake, the 87Sr/86Sr ratio is a function of surface water chemistry, and that water is fundamentally influenced by bedrock. Here is the interesting part of the story. The ratio can be used to determine whether uptake and growth occurred in geologically distinct areas with unique 87Sr/86Sr ratios, which is the case in the Mohawk. The signal can show when organisms spent time in isotopically distinct areas, thus it has been used in understanding the migration of birds, fish, and people (see Crowley et al., 2017). But the otolith in a fish is spectacularly suited to this sort of chemical chronology because it constantly accretes and is essentially in equilibrium with ambient water.
If an area has a distinctive Sr signature, one can develop a “strontium isoscape” where distinctive Sr ratios in water reflect bedrock Sr values (Crowley et al., 2017). If Sr isoscapes can be established, then that fingerprint can be used to understand the migration and movement of organisms, such as fish. In the Appalachians and this part of NY, limestone bedrock has low 87Sr/86Sr ratios, but granites and many metamorphic rocks have high 87Sr/86Sr ratios. From a geological perspective, the Mohawk has a more non-radiogenic signature and the upper Hudson has a more radiogenic signature.
Strontium fingerprint in the Mohawk
Blueback Herring from the Mohawk are different from peers in other rivers according to the Turner and Limburg study (2014). They show that the Mohawk Blueback Herrings have the highest Sr/Ca ratio, the lowest Ba/Ca ratio, and the lowest 87Sr/86Sr ratio, making them distinctive in general and certainly distinctive from those of the upper Hudson River (see their Table 3).
A combination of distinct geologies and use of different nursery habitats drove the strong differences in otolith signatures of river herring over relatively small spatial scales. Geological differences between the Adirondack Mountains and the Mohawk Valley drove the differences in otolith 87/86 Sr of Blueback Herring from the Mohawk River and [upper Hudson watershed] (p. 1540 Turner and Limburg, 2014).
The Sr fingerprint allowed a determination that Blueback Herring from the Mohawk River make a significant contribution to the Hudson River spawning stock (Turner and Limburg, 2014 – p. 1542). Thus a key question is the migration of this isotopically and geochemically distinct fish born in the Mohawk River.
The isotopic ratio in growth rings in otoliths show in incredible detail the changes in water chemistry over time in migrating fish that originated in the Mohawk (Limburg and Turner, 2016). These remarkable results reveal that some fish start in the Mohawk, migrate through the Hudson, went out into the ocean, and then returned. This pattern was revealed by the interesting isotopic and geochemistry of otolith profiles.
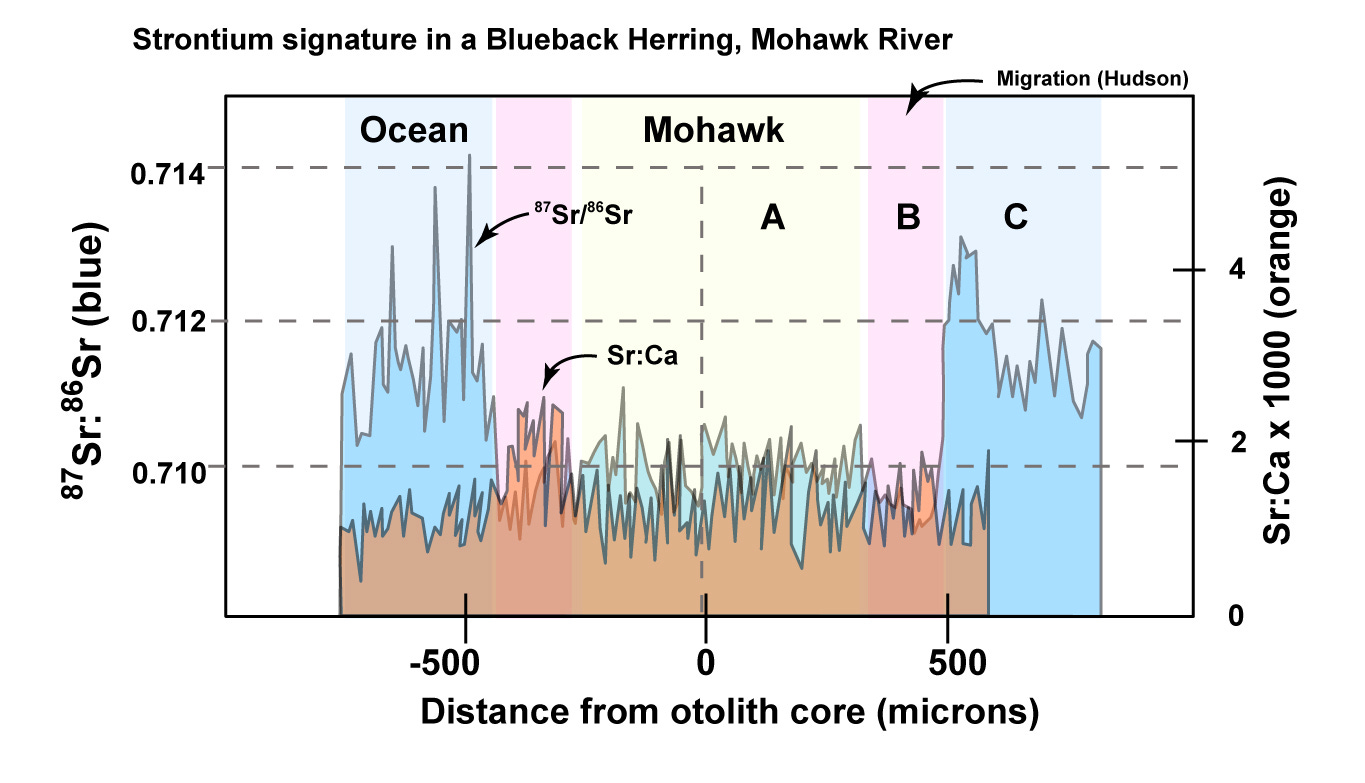
Diagram modified from Limburg and Turner, 2016 showing the distinctive strontium fingerprint in a otolith from a single Mohawk-born Blueback herring. 87Sr/86Sr results (blue, left scale) and Sr:Ca (orange, right scale) are here plotted from either side of the center of the otolith (0 microns - or birth of the fish). The fish was born in the Mohawk (A), migrated through the Hudson (B), and went to the Atlantic (C). It was three years old when captured (see Limburg and Turner, 2016 – Figure 3). The life story of a herring is captured in strontium isotopes.
This work shows that otoliths in Mohawk River Herring are distinctive from those of the Hudson. The 87Sr/86Sr ratio of first-year herring is 0.7107 in the Hudson and 0.7094 in the Mohawk: both are very much lower than ocean water. In addition, the Sr/Ca ratio is higher in the Mohawk than in the Hudson (the non-marine or non-estuary part). The Sr/Ca ratio is ~0.0016 in the Mohawk and 0.0010 to 0.0013 in the Hudson River (Limburg and Turner, 2016). Combining the two proxies allows for a distinctive otolith tag for Mohawk fish.
This stunning result means that the imprint on the fish otolith reflects the difference in the geology between the Mohawk and the Hudson. The Mohawk is underlain by a greater fraction of Cambrian and Ordovician limestones, which are mainly the Beekmantown and Trenton groups that are c. 450-500 Ma. These limestones are widespread in the Mohawk Valley, and thus they would provide both strontium and also a lower 87Sr/86Sr ratio because they tend to have a low fixed ratio.
Recall that River Herring are anadromous fish that are a cornerstone species. Understanding migration patterns in and out of the Mohawk River is important. Recall too that this annual spring migration relies on the operation of the locks at the Waterford Flight. It seems remarkable that the fish have figured out this passage avenue, but they have had this option for nearly 200 years. An important finding from the Turner and Limburg study (2014) is that the fraction of River Herring caught in the Hudson but born in the Mohawk may be as high as 30%. They note:
The contribution of Mohawk River Blueback Herring to the Hudson River population has important management implications, as fish can only enter the Mohawk River when the locks begin operating (typically on May 1). Because climate changes have led to earlier and more rapid warming of temperatures in the spring, the Mohawk River subpopulation may be negatively impacted if lock operation is not adjusted to allow earlier immigration.
Do we need to start operating the Locks on the lower part of the Mohawk River to facilitate fish passage to sustain these critical cornerstone species? Perhaps, but consider the fact that Blueback Herring were not present in the Mohawk prior to establishment of the locks. They are an invasive species.
Thus we end with a dilemma because the Mohawk now serves as a refuge for this otherwise stressed species. On one level, facilitating fish passage by operational changes to the Locks would seem to the the right thing to do.
This and other Notes from a Watershed are available at: https://mohawk.substack.com/,
Further reading
Campana, S.E., 1999. Chemistry and composition of fish otoliths: pathways, mechanisms and applications. Marine Ecology Progress Series, 188, pp.263-297.
Crowley, B.E., Miller, J.H. and Bataille, C.P., 2017. Strontium isotopes (87Sr/86Sr) in terrestrial ecological and palaeoecological research: empirical efforts and recent advances in continental‐scale models. Biological Reviews, 92(1), pp.43-59.
Limburg, K.E. and Turner, S.M., 2016. How common is “non-textbook” migration in Hudson River Blueback Herring?. Estuaries and Coasts, 39(4), pp.1262-1270.
Turner, S.M. and Limburg, K.E., 2014. Determination of river herring natal origin using otolith chemical markers: accuracy as a function of spatial scale and choice of markers. Transactions of the American Fisheries Society, 143(6), pp.1530-1543.